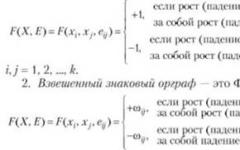Московский Государственный Университет имени М. В. Ломоносова
Биологический факультет, кафедра антропологии
Москва, 2001
Введение
Популяционная генетика – наука о генетических основах популяции. Это теоретическая область генетики, изучающая наследственную преемственность в популяциях, занимающаяся генетическим описанием и математическим исследованием самих популяций, и сил, на них действующих.
Наследственные изменения, происходящие в ряду поколений, лежат в основе процесса эволюции, поэтому популяционную генетику можно рассматривать как основу синтетической теории эволюции, т.е. объединения дарвиновской идеи естественного отбора и корпускулярной теории Менделя. Математическая основа популяционной генетики была доказана Рональдом А. Фишером (Fisher, 1930) в его фундаментальной теории естественного отбора.
Большая панмиксная популяция
Популяция - это сообщество особей одного вида, обладающих общим ареалом в течении длительного времени, расположенное отдельно от других сообществ и свободно скрещивающиеся между собой. Родственными узами связаны члены любой популяции, однако у организмов, размножающихся бесполым путем, отсутствуют связи, возникающие в результате перекрестного оплодотворения. Сообщество особей, размножающихся половым путем называется менделевской популяцией.
Причиной того, что отдельно взятый генотип не может служить адекватной еденицей процесса эволюции, является то, что его генотип остается неизменным на протяжении всей его жизни, а время жизни организма ограничено. С другой стороны, популяция представляет собой непрерывный ряд поколений, а генетическая структура популяции может изменяться, т.е. эволюционировать, от поколения к поколению. Непрерывность существования популяции обеспечивается механизмом биологической наследственности.
При изучении процесса эволюции важное значение имеет представление о генофонде. Генофондом называется совокупность генотипов всех особей популяции. Для диплоидных организмов генофонд популяции, насчитывает N особей, состоящих их 2N гаплоидных генотипов. Таким образом, генофонд популяции из N особей включает в себя N пар гомологичных хромосом и 2N аллелей каждого локуса (4N, т.к. у диплоидного организма 2 локуса). Исключение составляют половые хромосомы и сцепленные с полом гены, представленные в каждом гетерогаметном организме в одном экземпляре.
Непосредственно мы наблюдаем лишь фенотипы, а не генотипы или гены. Изменчивость генофонда может быть описана либо частотами генов, либо частотами генотипов. Если мы знаем соотношение между генотипами и соответствующими им фенотипами, то по частотам наблюдаемых фенотипов мы можем рассчитать частоты соответствующих генотипов.
Частоты аллелей можно рассчитать по частотам генотипов, учитывая, что в гомозиготах содержится по два одинаковых аллеля, а в гетерозиготах – по одному аллелю каждого типа. Таким образом, что бы получить частоту аллелей каждого типа, нужно к частоте индивидуумов, гомозиготных по данному аллелю, прибавить половину частоты гетерозигот по этому аллелю. Если частоты генотипов представить как: гомозиготных (АА) – D, (аа) – R, гетерозиготного (Аа) – H, то частоты аллелей считаются как:
Одна из причин, по которым генетическую изменчивость популяций часто предпочтительнее описывать, используя частоты аллелей, а не генотипов, состоит в том, что различных аллелей обычно бывает гораздо меньше, чем генотипов. При двух аллелях число возможных генотипов равно трем, при трех аллелях – шести, при четырех – десяти. В общем случае, если число различных аллелей одного локуса равно k, то число возможных генотипов равно k(k + 1)/2.
В 40-е – 50-е годы существовало две конкурирующих гипотизы о генетической структуре природных популяций. Согласно классической модели, генетическая изменчивость популяции очень мала, а согласно балансовой – очень велика.
По классической гипотизе подавляющее большинство локусов содержит аллели так называемого "дикого типа" с частотой, близкой к единицы. Типичная особь гомозиготна по аллелю "дикого типа" или гетерозиготна по мутантному аллелю и аллелю "дикого типа". Нормальный ("идеальный") генотип гомозиготен по аллелям "дикого типа" . Согласно балансовой теории, не существует какого-то одного аллеля "дикого типа", а присутствует целый ряд аллелей с различными частотами, следовательно, популяцию составляют особи, гетерозиготные по этим аллелям. Нормальный генотип отсутствует.
Биологическая эволюция – это процесс накопления изменений в организме и увеличения их разнообразия во времени. В своей основе эволюционные изменения имеют генетический аспект, т.е. происходит изменение наследственного вещества, которое, при взаимодействии со средой, определяет все признаки организма. На генетическом уровне эволюция представляет собой накопление изменений в генетической структуре популяций.
Эволюцию можно рассматривать как двуступенчатый процесс. С одной стороны. возникают мутации и рекомбинации – процессы, обуславливающие генетическую изменчивость; с другой стороны, наблюдается дрейф генов и естественный отбор – процессы, посредством которых генетические изменения передаются из поколения в поколение.
Эволюция возможна только в том случае, если существует наследственная изменчивость. Единственным поставщиком новых генетических вариантов служит мутационный процесс. Помимо мутаций к процессам, изменяющим частоты аллелей в популяции, относят естественный отбор, поток генов (т.е. миграция их) между популяциями и случайный дрейф генов. Частоты генотипов (но не аллелей!) могут изменяться также в результате ассортотивного, т.е. неслучайного, формирования брачных пар.
Итак, для описания генетической структуры популяций надо знать число аллелей и генотипов. Пусть генотипы различаются фенотипически: АА, Аа, аа; их частоты равны соответственно: D, H, R. Пусть p – частота аллеля А, q – частота аллеля а. В популяции, содержащей диплоидных особей, частоты аллелей будут равны:
Наследственность сама по себе не изменяет частот генов. Этот принцип известен под названием закона Харди-Вайнберга, фундаментального закона популяционной генетики.
Основное утверждение закона состоит в том, что в отсутствии элементарных эволюционных процессов, а именно мутаций, миграций, отбора и дрейфа генов, частоты генов остаются неизменными из поколения в поколение. Этот закон утверждает также, что если скрещивание случайно, то частоты генотипов связаны с частотами генов простыми (квадратичными) соотношениями. Случайное скрещивание происходит тогда, когда вероятность формирования брачной пары между особями не зависит от их генетической конституции. Когда на выбор брачного партнера оказывает влияние генотип, говорят об ассортативном скрещивании. Из закона Харди-Вайнберга вытекает следующий вывод: если частоты аллелей у ♂ и ♀ исходно одинаковы, то при случайном скрещивании равновесные частоты генотипов в любом локусе достигается за одно поколение.
Закон Харди-Вайнберга сформулировали в 1908 году независимо друг от друга математик Т. Харди (Англия) и врач В.Вайнберг (Германия). Он гласит, что процесс наследственной преемственности не влияет на число частот аллелей и (при случайном скрещивании) генотипов по определенному локусу. При случайном скрещивании равновестные частоты генотипов по данному локусу достигаются за одно поколение, если исходные частоты аллелей одинаковы у обоих полов.
Равновесные частоты генотипов задаются произведением частот соответствующих аллелей. Если имеется два аллеля, А и а, с частотами p и q, то частоты трех возможных генотипов выражаются уравнением
(p + q)2 = p2 + 2pq + q2 = p2 + 2pq + q2, где p2 = AA = D, 2pq = Aa = H, q2 = aa = R.
С учетом того, что сумма всех частот аллелей, как и сумма всех частот генотипов, всегда равна единице, то (p + q)2 = 1. Рассмотрим различные типы браков и их постоянства в двуаллельной популяции.
Таблица 1
| тип брака | частота | потомство | ||
| АА | Аа | аа | ||
| АА x АА | D2 | D2 | ||
| АА x Аа | 2DH | DH | DH | |
| АА x аа | 2DR | 2DR | ||
| Аа x Аа | H2 | 1/4H2 | 1/2H2 | 1/4H2 |
| Аа x аа | 2RH | RH | RH | |
| аа x аа | R2 | R2 | ||
| D+H+R | (D+1/2H)2 | 2(D+1/2H)(R+1/2H) | (R+1/2H)2 | |
| сумма | 1 | p2 | 2pq | q2 |
Получаем, что в большой панмиксной популяции не зависимо от исходных генетических структур в первом же поколении после случайного скрещивания достигаются равновестные частоты генотипов.
Если у ♂ и ♀ генотипы разные, а численность полов одинакова, то для достижения равновесия потребуется два поколения
| ♂ | ♀ | |||
| AA | x | aa | ||
| Аа | x | Аа | ||
| 1/2AA | Aa | 1/2aa | ||
| 1/4 | : | 1/2 | : | 1/4 |
| 1 | : | 2 | : | 1 |
Одно из возможных применений закона Харди-Вайнберга состоит в том, что он позволяет рассчитать некоторые из частот генов и генотипов в случае, когда не все генотипы могут быть идентифицированности вследствии доминантности и гетерозиготности некоторых аллелей. Из закона Харди-Вайнберга можно вывести интересное следствие: разные аллели присутствуют в популяции главным образом в гетерозиготном, а не в гомозиготном состоянии. Рассмотрим пример. Альбинизм человека обусловлен довольно редким рецессивным геном. Если аллель нормальной пигментации обозначить как А, а аллель альбинизма – а, то генотип альбиносов будет аа, генотип нормально пигментированных людей – АА и Аа. Предположим, что частота генотипов альбиносов (частота рецессивной гомозиготы) равна 0,0001, т.е. q2 = 0,0001, тогда. q = 0,01, соответственно р = 1 – 0,01 = 0,99; частота генотипа АА будет равна 0,9801, а генотипа Аа – 2pq = 0,02. Следовательно, в гетерозиготном состоянии находится примерно в 100 раз больше рецессивных аллелей, чем в гомозиготном.
Для генов, сцепленных с полом, равновесные частоты генотипов у ♀ (т.е. у гомогаметного пола) совпадают с равновесными частотами аутосомных генов (p2 – AA, 2pq – Aa, q2 – aa). Частоты гамет гемизиготных ♂ (т.е. гетерогаметного пола) совпадают с частотами аллелей: p – А, q – а. Из этого следует, что фенотипы, определяемые рецессивными генами, у ♂ встречаются чаще, чем у ♀. Этим объясняется то, что наследственным заболеваниям, вызываемыми рецессивными, сцепленными с полом генами, больше подвержены ♂.
Генетическое представление об изменчивости
Закон Харди-Вайнберга гласит, что при отсутствии возмущающих процессов частоты генотипов не изменяются. Однако процессы, изменяющие частоты генов, постоянно происходят в популяции, и без них не было бы эволюции.
Рассмотрим процесс мутирования. Мутации – спонтанные изменения, происходящие в ДНК. Они бывают точечные, или генные (самые локализованные мутации, происходят в самом гене), хромосомные (изменяющие структуру хромосом, например, делеции, транслокации) и геномные (изменяющие число хромосом). Мутации вносят в популяцию новые аллели. Это чрезвычайно медленный процесс, т.к. сами по себе они изменяют генетическую структуру популяции с очень низкой скоростью. Если бы мутации были единственным процессом, обуславливающим эволюционные изменения в популяциях, то эволюция протекала бы очень медленно.
Направленность мутаций в природе случайна и непредсказуема. Каким образом они появляются в популяциях и как распространяются? Рассмотрим пример.
В популяции, целиком состоящей из особей с генотипом АА, возникает мутация и появляется одна гетерозигота Аа. Предположим, что а эволюционно нейтральна (не подвержена действию естественного отбора). Гетерозигота АА скрещивается с гомозиготой аа:
потомки (в отношении) АА: Аа (1/2: 1/2),
т.е. ген а может остаться в популяции или исчезнуть с вероятностью 1/2. Если он останется и снова произойдет скрещивание, то во втором поколении новая аллель исчезнет из популяции или удвоится с вероятностью 1/4 и т.д. Вероятность сохранения/удаления из популяции мутации зависит от числа потомков и находится как (1/2)k, где k – число потомков, k = 0, 1, 2, ...
Исспользуя уравнение распределения Пуассона, можем рассчитать процент вероятности сохранения и потери нейтральной мутации. Для первого поколения вероятность сохранения мутации (в %) равен 63, потери –37, для второго соответственно 47 и 53, для третьего – 63 и 37 и т.д. Предельная вероятность исчезновения мутации равна 100%; в 127 поколении вероятность исчезновения мутации равна 98, 47% (почти 100%), следовательно, мутация исчезнет из популяции приблизительно через 130 поколений. Потеря мутаций – необратимый процесс.
Рассмотрим теперь повторные мутации. Скорость (или частота) возникновения мутаций является отношением числа мутировавших аллелей к общему числу аллелей в популяции. Она зависит от типа аллеля, а также от внутренних и внешних условий, меняется под действием различных факторов, например, радиации, высоких температур и т.п. При естественных условиях скорость возникновения мутаций колеблется от 10(-4) –10(-7).
Пусть в популяции нормальная аллель А с частотой в начальный момент времени равной p0, а мутантный аллель – а(q0). Скорость мутации из А в а – U. Общая формула частот аллеля А будет pn = p0(1–U)n, а аллеля а – qn = 1 – p0(1 – U)n. Изменение частоты мутаций за поколение: Δq = Upn, т.е. чем больше р, тем с большей частотой аллель А переходит в аллель а.
Мутации могут происходить и в обратном направлении, за счет чего происходит компенсация и сохранение генетического разнообразия. Если А мутирует в а со скоростью U, а а обратно мутирует в А со скоростью V, тогда равновесные частоты аллелей будут равны
p* = V/(U + V), q* = U/ (U + V).
Отметим два обстоятельства. Во-первых, частота аллелей обычно не находится в состоянии, отвечающим равновесию между прямыми и обратными мутациями, поэтому на них влияют и другие процессы (например, естественный отбор). Во-вторых, при наличии прямых и обратных мутаций изменения частот аллелей происходят медленее, чем в том случае, когда мутации идут только в одном направлении, посколько обратные мутации частично компенсируют изменение частоты аллелей в результате прямых мутаций. Это еще раз доказывает, что для того, что бы мутации сами по себе привели к сколько-нибудь значительному изменению частот аллелей, требуется много времени.
Естественный отбор
К идее естественного отбора как основного процесса эволюции пришли независимо друг от друга Чарльз Дарвин и Альфред Рассел Уоллес. В 1858 году на заседании Линнеевского общества в Лондоне были представлены сообщения об их открытии. Доказательства того, что эволюция происходит именно путем естественного отбора, были представлены Дарвином с приведением множества примеров в его работе "Происхождение видов", опубликованной в 1859 году.
Основой теории естественного отбора является факт, что некоторые генотипы в популяции имеют преимущества перед другими, как в выживании, так и при репродукции. То, что генотипы в данных условиях имеют возможность выжить и оставить плодовитое потомство, называется генетической приспособленностью. Она обуславливает направленное сохранение генотипа.
Естественный отбор благоприятствует (и противодействует) сохранению генотипа только через посредство фенотипа. В связи с этим выделено два исхода действия отбора: положительный отбор (сохранение "полезных" генотипов) и отрицательный отбор (эллиминация "вредных" генотипов). Таким образом, естественный отбор – это направленный процесс, движущая сила эволюции.
В качестве количественной меры отбора используется относительная приспособленность (называемая также селективным, или адаптивным значением). Приспособленность является мерой эффективности размножения данного генотипа, мерой вклада его в следующее поколение.
Особенности существования организма на различных ступенях жизненого цикла могут оказывать влияние на его репродуктивный успех, определяющий направление естественного отбора и, следовательно, на приспособленности генотипов. Эти особенности сказываются на выживаемости, скорости развития, успешности спаривания, плодовитости и т.п., т.е. на величинах, называемых компонентами приспособленности. Важнейшими компонентами являются выживаемость и плодовитость. Другие компоненты могут рассматриваться самостоятельно или включаться в две основные.
Математические модели естественного отбора были описаны Райтом и Фишером. Простейшая модель – популяция с одной парой аллелей, приспособленность к которой не зависит от других локусов.
Пусть АА, Аа, аа – генотипы популяции, их относительные приспособленности соответственно W11, W12, W22. Частоты генов равны p и q.
Таблица 2*
* – Т – сумма частот генов после отбора, p1,q1 – новые частоты генов, p1=(p2W11+pqW12)/T, q1=(q2W22+pqW12)/T.
Рассмотрим модели действия естественого отбора.
1. Полная эллиминация рецессивных гомозигот (летальных).
Генетическая приспособленность рецессивных гомозигот равна нулю, т.к. они летальны (либо не доживают до репродуктивного возраста, либо не оставляют потомства). Примером служит болезнь фенилкетонурия.
Таблица 3*
* – частоты аллелей после отбора: p1=1/(1+q); q1=q2/(1+q).
Количество изменений аллелей за одно поколение будет: Δq=–q2/(1+q). Δq пропорциональна q2 (частоте рецессивных гомозигот), т.е. чем больше частота, тем больше количество изменений. Δq всегда отрицательная величина (или равная нулю), значит значение q уменьшается в результате отбора. Иногда происходит не полная эллиминация, а частичный отбор, т.е. не все особи доживают до репродуктивного возраста и оставляют жизнеспособное потомство.
Введем понятие коэффициент отбора S, W = 1 – S. S пропорциональна снижению воспроизводства генотипа по сравнению с нейтральным, приспособленность которого условно принимается за единицу. Для нейтральных признаков S = 1, для нейтральных – S = 0.
Таблица 4*
* – частоты аллелей после отбора: p1= p/(1–Sq2); q1=q2(1–Sq)/(1–Sq2).
Примером отбора против рецессивных гомозигот служит явление индустриального меланизма, изученное в Англии на бабочках Bistonbetularia. До середины XIX века эти бабочки имели светло-серую окраску. Затем, в промышленных районах, там, где стволы деревьев постепенно почернели от копоти и сажи, стали появляться темные бабочки. В некоторых местностях темная разновидность почти полностью вытеснила светлую. Светло-серые бабочки гомозиготны по рецессивному аллелю, темные – либо гетерозиготны, либо гомозиготны по доминантному аллелю.
Вытеснение в промышленных районах светлой разновидности Bistonbetularia темной происходило благодаря избирательному истреблению бабочек птицами: на почерневшей от копоти коре светлые бабочки становились более заметными, в то время как темные были хорошо замаскированы. Бабочек отлавливали, метили и при повторном отлове доля темных бабочек составляла 53%, а светлых – 25%. Посколько плодовитость обеих форм примерно одинакова, можно предположить, что их относительные приспособленности определяются исключительно в выживаемости, обусловленной их неодинаковой уязвимостью для насекомоядных птиц.
2. Отбор против доминантных аллелей.
Это часто встречающийся тип отбора, к нему относятся почти все геномные и доминантные генные мутации.
Отбор против доминантных аллелей идет более эффективно, чем отбор против рецессивных, поскольку доминантные аллели проявляются не только в гомозиготном, но и в гетерозиготном состоянии.
Таблица 5*
* – частоты аллелей после отбора: p1= p(1–S)/T; q1=q(1–pS)/T.
Изменение частоты доминантного аллеля: p=–(Sq2(1–q))/(1–Sq2), уже за одно поколение отбора частота p уменьшится. Если доминантная аллель летальна, то Δp = –p и за одно поколение аллель полностью исчезнет из популяции.
3. Отбор в пользу гетерозигот.
Такой отбор происходит, когда обе гомозиготы имеют пониженную приспособленность по сравнению с гетерозиготой (W11
Таблица 6*
*– частотыаллелейпослеотбора: p1= (p– p2S1)/(1–S1 p2–S2q2); q1= (q – q2 S2)/(1–S1 p2–S2q2).
Δq=pq(pS1– qS2)/(1–S1 p2–S2q2); при положительных значениях частота рецессивного признака увеличивается, при отрицательных – уменьшается, до тех пор. пока не достигнется состояние равновесия, т.е. pS1=qS2. Равновестные частоты равны q*= S1/(S1+ S2), p*=S2/(S1+ S2). Равновесия при отборе в пользу гетерозигот – устойчивое, оно определяется коэффициентом отбора.
Хорошо известным примером сверхдоминирования может служить серповидноклеточная анемия, широко распространенная в некоторых странах Африки и Азии. Нормальный гемоглобин обозначается HbAHbA, аномальный – HbSHbS. Возможно наличие трех вариантов генотипов: HbAHbA (1– S1), HbAHbS (1), HbSHbS (1–S2). S2 близок к единице, т.к. HbSHbS редко выживают. Приспособленность HbAHbA близка к единице в районах, где малярия не наблюдается. Из этого следует, что q*= S1/(S1+ S2) приблизительно равна S1/(1+S1).
Серповидноклеточная анемия представляет собой пример зависимости приспособленности генотипов от окружающей среды. В тех местах, где малярию искоренили или где ее никогда не было, гомозиготы HbAHbA обладают одинаковой приспособленностью с гетерозиготами. При этом направление отбора изменяется, он уже не благоприятсвует гетерозиготам, а направлен против рецессивных гомозигот и приводит к эллиминации рецессивного аллеля.
4. Отбор против гетерозигот.
Возможны ситуации, в которых гетерозигота обладает более низкой приспособленностью, чем обе гомозиготы. Примером такого рода могут служить транслокации, что связано с более низкой плодовитостью гетерозигот в данном случае.
Таблица 7*
*– частоты аллелей после отбора: p1= p(1– pS)/(1–2Spq); q1= q(1– pS)/(1–2Spq).
Δq=2Spq(q–1/2), 1/2 – точка равновесия, равновесие неустойчивое.
5. Частотно – зависимый отбор.
К устойчивому генетическому полиморфизму может приводить не только преимущество гетерозигот, но и частично-зависимый отбор, при котором приспособленности генотипов изменяются в зависимости от их частот. Предположим, что приспособленности двух генотипов, AA и aa, связаны с их частотами обратной зависимостью: приспособленность велика, когда генотип редок, приспособленность мала, если генотип широко встречается в популяции. При изменчивой среде редким генотипам может быть свойственна высокая приспособленность, поскольку сочетание условий, при которых отбор благоприятствует таким генотипам, могут встречаться довольно часто. Когда же какой-нибудь генотип широко распространен в популяции, он может обладать низкой приспособленностью, т.к. благоприятствующие ему сочетания внешних условий будут встречаться значительно реже.
Частотно-зависимый половой отбор возникает, когда вероятность скрещиваний зависит от частот соответствующих генотипов. Нередко при выборе брачных партнеров предпочтение придается носителям редких генотипов, особенно если они выражены фенотипически. Это предпочтение представляет собой один из механизмов поддержания генетического полиморфизма в популяциях, поскольку приспособленность генотипа повышается по мере того, как он становится редким. Частотно-зависимый отбор особенно важен при наличии миграций.
6. Общая модель действия отбора на популяцию по одному локусу.
Эта модель связана с понятием средней приспособленности популяции, Wср.
Таблица 8*
* – Wср= W1 p2+2W2pq+ W3 q2.
Общая формула для всех типов отбора следующая:
Δq=(pqdWср)/(2Wсрdq); при равновесии Δq=0, dWср/dq=0, между средней приспособленностью и изменением частоты рецессивного аллеля наблюдается обратная пропорциональность.
Совместное действие мутаций и отбора
Во всех случаях отбора его окончательный результат был одним и тем же – "вредный аллель" полностью эллиминировался из популяции. Присутствие этих аллелей в популяции поддерживалось мутациями. Эффекты этих двух процессов уравновешивали друг друга, когда число вредных аллелей, исчезнувших и появившихся в результате двух этих процессов, было одинаково.
В равновесном состоянии между действием мутаций и отбора против рецессивных гомозигот, при U – скорости мутирования за поколение и S – интенсивности отбора, частота рецессивного аллеля q* =(U/S)1/2, а для летальных рецессивных гомозигот q* =(U)1/2.
Равновесие устойчивое, благодаря чему "вредные" гены нельзя удалить из популяции.
Для случая, когда происходят мутации и отбор против доминантных аллелей, скорость мутирования равна V, p* =(V/S)1/2, для летали – p* =(V)1/2. Пример – брахидактилия.
Случай неустойчивого равновесия наблюдается при отборе против гетерозигот, W1=W2, q* =U/S.
Миграция и инбридинг
Миграции, или поток генов, возникает, когда особи из одной популяции перемещаются в другую и скрещиваются с представителями второй популяции. Поток генов не изменяет частот аллелей у вида в целом, однако в локальных группах они могут изменяться. Эффективность обмена генами зависит от структуры популяции, от интенсивности миграции и различий в частотах гена.
Если в локальную популяцию с определенной частотой входят мигранты (скорость миграции – m), то в следующем поколении потомки получат долю генов m от мигрантов и (1–m) – от аборигенов. С учетом начальных частот генов q0 и Q соответственно для исходной и соседней популяций, получим изменение гена за одно поколение Δ q= – m(q0– Q) и через n поколений qn–Q =(1– m)n(q0– Q); частоты генов локальной популяции будут стремиться к частотам гена мигрантов.
Интересную форму ассортативного скрещивания представляет собой инбридинг, когда скрещивание между родственными особями происходит чаще, чем можно было бы ожидать на основе случайного скрещивания. Ограниченность в выборе партнера приводит к тому, что у одной особи могут встретиться два идентичных аллельных гена, имеющих общее происхождения.
Самым крайним случаем инбридинга является самооплодотворение, или самоопыление. Инбридинг часто используют в садоводстве и животноводстве. В человеческих популяциях инбридинг повышает частоту проявления вредных рецессивных аллелей.
Вероятность инбридинга описывает коэффициент инбридинга (F), являющийся мерой его генетических последствий. Вычислить его можно, опираясь на частоты аллелей родителей и потомства, а также при помощи так называемого коэффициента путей, для нахождения которого строится родословная интересующих нас особей, выбирается один из ближайших предков, для которого существует R путей, состоящих из шагов: m1, m2,..., mx. Для t общих предков: R=1/2(∑ по i от 1 до t) x. Отсюда F=1/2(∑ по i от 1 до t)((1/2)mi–1).
Приведем примеры коэффициента инбридинга F для различных случаев скрещивания:
F(дядя\тетя x племянница\племянник)= 1/8
F(двоюродные сибсы)= 1/16
F(двоюродный(-ая) дядя\тетя x племянница\племянник)= 1/32
F(троюродные сибсы)=1/64
F отражает в популяции избыток особей, гомозиготных по какому-нибудь локусу, показывая также увеличение доли гомозиготных локусов в генотипах отдельных особей.
Учитывая коэффициент инбридинга, мы можем получить обобщенный закон Харди-Вайнберга, называемый законом Райта.
Пусть в гомозиготе AA с вероятностью F оба аллеля идентичны, а с вероятностью (A–F) – независимы, т.е. происходят от разных предковых аллелей. Частота AA равна p, частота aa – q; с вероятностью pF гомозигота AA будет идентична, с вероятностью qF гомозигота aa будет идентична.
Частоты генотипов в популяции (при инбридинге) будут следующие:
Отсюда видно, что частота гомозигот увеличивается, а частота гетерозигот уменьшается при имбридинге. При этом коэффициент имбридинга Райта (F) не связан с изменением частот аллелей, а указывает на то, как аллели объединяются в пары.
Имбридинг обычно приводит к понижению приспособленности потомства вследствии ухудшения таких важных характеристик организма, как плодовитость, жизнеспособность, устойчивость к болезням. Это явление принято называть имбредной депрессией. Она обусловлена повышением степени гомозиготности по вредным рецессивным аллелям. Имбредной депрессии можно противопоставить скрещивание между представителями независимых имбредных линии. Такие гибриды обычно обнаруживают заметно возросшую приспособленность. Говорят, что такие гибриды обладают гибридной мощностью, или гетерозисной силой. Независимые имбредные линии обычно становятся гомозиготными по "вредным" аллелям; при скрещивании между двумя такими линиями можно сохранить в потомстве гомозиготность по искусственно отобранным признакам, тогда как "вредные" аллели переводятся в гетерозиготное состояние.
У человека имбридинг приводит к возникновению имбредной депрессии, поэтому в большинстве человеческих культур запрещены кровнородственные браки, хотя в некоторых популяциях они все же существуют. С учетом накопления вредных аллелей в потомстве от браков между родственниками, частота появления новорожденных с различными дефектами примерно вдвое выше, чем в потомстве не состоящих в родстве супругов. Впрочем, это касается рецессивно наследуемых дефектов; вероятность появления доминантнозависимых дефектов от родственных браков не выше, чем в потомстве от неродственных браков.
Генетическая структура подразделенной популяции
Большая популяция называется подразделенной, если она состоит из некоторого числа популяций, каждая из которых является самостоятельной единицей, скрещивание между ними происходит случайно (в зависимости от типа изоляции – полной или частичной).
Случай полной изоляции.
Пусть частота аллеля a – qi в i-группе, рi – частота A, k – количество групп. qi + рi=1. Среднее значение частот по группам qср и рср:
qср= (∑qi)/k
рср=(∑ рi)/k
Насколько велика изменчивость гетерогенной популяции, можно узнать с помощью Ω2 (вариансы):
Ω2q= Ω2p= (∑qi2)/k – qср2=(∑ pi2)/k – pср2
AA – pср2+ Ω2 рi
Aa=2piqi–2Ω2 q2
aa – qср2+ Ω2 qi
Получаем. что процент гомозигот увеличивается (как при имбридинге).
Случай частичной изоляции (последствия миграций).
При миграциях различия между группами сглаживаются, Ω2 (и F) уменьшаются. Этот случай хорошо иллюстрирует островная модель Райта.
qi!= qi – m(qi –qср)
Δ q= – m(qi –qср)
Ω2 qi= (1–m)2Ω2 q2
Частоту мутаций определяют двумя методами.
Прямой метод. Применяним только к доминантным мутациям для одного аллеля, признак которых не вызывается негенетическими факторами. Производится подсчет всех носителей доминантных мутаций в популяции, родители которых имели нормальные аллели и считается их отношения.
Косвенный метод. Подходит как для доминантных, так и для рецессивных мутаций (в гетерозиготе мутации не проявляются).
Допустим, что между подгруппами возможны миграции и на популяцию действует отбор, тогда для генотипов AA, Aa, aa, с промежуточными приспособленностями W1, W2, W3, равными соответственно 1, 1–S,
1–2S (S – коэффициент отбора), скорость миграции равна m.
q*= {(m+S)±((m+S)2–4mS qср)1/2}/2S.
Если S>0, то отбор будет направлен против рецессивных гомозигот, если S<0, то отбор будет им благоприятствовать. В зависимости от отношения m к S, новые частоты аллелей будут формироваться за счет отбора (|S|>>m) или миграций (|S|< Стохастические процессы в популяциях
Верное представление о численности популяции дает понятие ее генетической численности (Nэф=Ne), которая определяется по числу особей, дающих начало следующему поколению, а не общее число особей в популяции. Это объясняется тем, что в генофонд следующего поколение вносят вклад лишь особи, являющееся в предыдущем поколении родителями, а не вся популяция в целом, т.е. если численности ♂ равны N0, а ♀ – N1, то Ne = 4N0N1/(N0 + N1). Периодически происходят уменьшения численности популяции, что может привести к случайному дрейфу генов – к изменению частот аллелей в ряду поколений, вызываемому случайными причинами. Дрейф генов относится к особому классу явлений, называемым ошибками выборки. Общее правило, применительно к живым организмам, состоит в том, что чем меньше число скрещивающихся особей в популяции, тем больше изменений, обусловленных дрейфом генов, будут претерпевать частоты аллелей. Ошибки со временем накапливаются, образуя в популяции кумулятивный эффект. Кумулятивный эффект происходит в результате накопления изменений в процессе дрейфа генов. Если популяция не слишком мала, то обусловленные дрейфом генов изменения частот аллеля, происходящее за одно поколение, малы, но могут стать значительными, если остануться в популяции в ряду поколений. Если на частоты аллелей не оказывает влияние другие гены, то в конце концов один аллель зафиксируется, а другой эллиминируется. Когда на популяцию действует только дрейф генов, то вероятность фиксации частоты будет максимальной; предельным случаем будет возникновение новой популяции, состоящей всего из нескольких особей. Этот процесс называется "эффектом основателя" (Эрнест Майер). Такая ситуация возможна на изолированных территориях – на океанских островах, в закрытых озерах, лесах. Новая популяция может резко отличаться от прародительской. При резком изменении внешних условий популяция попадает в стрессовое состояние, ее численность может резко сократиться, возможно, даже до уровня вымирания. Если популяция не исчезнет, то в ней произойдет изменение частот аллелей, появление новых их сочетаний. Новая, отличная от родительской, популяция прошла через "бутылочное горлышко". Такая ситуация происходила в первобытных племенах людей, когда после упадка появлялись новые сообщества, численность которых восстанавливалась за счет мигрантов. Этот эффект, возможно, имеет отношение к возникновению различий в частотах аллелей между популяциями современного человека, например, в группах крови AB0. Взаимодействие основных факторов популяционной изменчивости
В популяциях частоты аллелей могут меняться под действием внешних и внутренних факторов, но форма взаимодействия между ними остается постоянной, поэтому говорят о стационарном взаимодействии между частотами в популяции. Приведем пример. Если в случайной выборке измерить рост отцов и их сыновей, то полученные средние значения и отклонения от них будут одинаковыми и у отцов, и у сыновей. Стационарное распределение действует и на изолированные популяции. Рассмотрим распределение определенного локуса в островной модели Райта, т.е. среди большого числа полностью или частично разрозненных групп, подвергающихся постоянному давлению внешних и внутренних факторов. Здесь стационарное распределение подразумевает тот факт, что хотя в ряду поколений значение частот в каждой отдельной группе может изменяться, но в равновестном состоянии форма распределения по группам во всех поколениях будет одинаковой. При разных условиях форма кривой распределения будет различной. Она зависит от размера популяции и от давления, которое оказывает на нее внешние и внутренние факторы. Теория адаптивных полей Райта выражает изменение средней приспособленности Wср и показывает распределение популяций на топографической карте. Значения Wср образуют в пространстве поверхность, состоящей из возвышений и впадин, которые являются соответственно максимальными и минимальными значениями. В большой популяции при изменении условий происходит сдвиг пиков и впадин и они могут превращаться друг в друга. Похожая ситуация происходит и в средней по размеру популяции, а в малой максимум достигается редко. В самом выгодном положение находится подразделенная популяция, в которой близость к пику дает возможность его освоить и увеличить свою численность. Подразделенная популяция представляется как определенная система, обладающая качественно иными свойствами, которыми не обладают ее составляющие; в ней факторы стабильности и изменчивости сбалансированы. Действие популяционных факторов в человеческих популяциях
Мутационный процесс Геномные мутации (изменения в количестве хромосом). Причинами геномных мутаций являются нерасхождения хромосом в мейозе, утрата одной из хромосом в результате анафазного отставания, нарушения в митотическом цикле. Трисомии. Синдром Дауна (трисомия по 21 хромосоме). Риск появления таких больных тесно связан с возрастом родителей (для ♀ после 35-45 лет, для ♂ – после 50). Частота появления синдрома у новорожденных 1: 700. Для больных характерны аномалии лица (узкие глазные щели, эпикантус, косоглазие), умственная отсталость и физическая недоразвитость. Они очень подвержены инфекционным заболеваниям и аномалиям сердца, поэтому продолжительность жизни таких больных невелика. Синдром Патау (трисомия по 13 хромосоме). Встречается в десять раз реже, чем синдром Дауна, появление зависит от возраста матери и времени зачатия (пик приходится на осень-зиму). Больные редко доживают до одного года, для них характерны микроцефалия, волчья пасть, заячья губа. Синдром Эдвардса (трисомия по 18 хромосоме). Частота возникновения – 1: 7000, также зависит от возраста матери. Больные погибают в младенческом возрасте. Все остальные трисомии летальны; в клетках самоабортируемых плодов обнаружены трисомии по другим хромосомам. Нерасхождение половых хромосом. Это более мягкие нарушения, затрагивающие половое развитие, психические растройства при которых не столь тяжелы. При таких нарушениях часто происходит внутриутробная гибель эмбрионов, что является одним из механизмов отбора у человека. Синдром Кляйнфельтера (XXY). Частота встречаемости – 1: 700. Это мужчины с феминоподобной внешностью, высокого роста, половые органы недоразвиты (в связи с пониженным синтезом андрогенов), бесплодные, умственное развитие немного ниже нормы. Частота встречаемости – 1: 2500. Клинически практически не отличается от XXY, но уровень умственного развития еще ниже. Встречается редко, характеризуется сильным слабоумием. Частота встречаемости – 1: 1000. Это женщины, мало отличающиеся от нормальных, иногда с недоразвитыми яичниками и мягкой формой олигофрении. Синдром Тернера (X0). Частота встречаемости – 1: 2500. Женщины небольшого роста, с общей задержкой психического развития, уровень IQ – 80-90. Частота встречаемости – 1: 800. Мужчины высокого роста, неандертальского типа, уровень IQ – 80-90. В 1965 году была выдвинута теория "хромосомных убийств", по которой наличие такого генотипа (лишней Y хромосомы у ♂) говорило об антисоциальном поведении. Теория не получила подтверждения. Частичная моносомия Синдром "кошачьего крика" (частичная делеция 5P (короткого плеча) хромосомы). Синдром характеризуется аномалией внутренних органов, идиотией. Больные не доживают до репродуктивного возраста. Синдром 18P. Один из наиболее часто встречающихся синдромов. Характеризуется умственной отсталостью. Сходная симптоматика у 18Q (длинное плечо), 21Q синдромов. Хромосомные мутации. В основном встречаются инверсии хромосом, при которых возможно слабоумие, небольшие мутации фенотипически не проявляются. Генные мутации. Для генных мутаций характерна сильная вариабельность. В одних случаях мутации незаметны, в других вызывают гибель организма. Причинами большинства заболеваний являются изменения в биохимических реакциях организма (фенилкетонурия, болезнь Тея-Сакса). Распределение таких мутаций в разных популяциях неодинаково, оно отражает ее генетическую историю. Некоторые мутации встречаются только на определенных территориях, что часто связано с изоляционным фактором, например, цветовая слепота на одном из островов Меланезии, наследственная слепота на острове Тристан, альбинизм в Альпах. 2. Инбридинг
В изолятах и полуизолятах число близкородственных и кровнородственных браков гораздо выше, чем в неизолированных популяциях. Браки между лицами одной расы, религии повышают вес родственной связи. В большинстве стран кровнородственные браки запрещены, хотя иногда они имеют место. Браки между близкими родственниками были приняты в древности у представителей царских кровей в Египте, Индии и других странах. Сейчас под действием социальных факторов они практикуются в Японии, Индии. Существуют даже целые популяции с ярко выраженным генетическим грузом ("вредными" мутациями), например, цыгане Южного Уэльса, каждый четвертый из которых несет ген фенилкетонурии или жители деревни Новая Германия (Парагвай), жертвы евгеники. Инцестные браки повышают частоту проявления "вредных" рецессивных аллелей, что приводит к большому количеству серьезных нарушений. 3. Естественный отбор
За время своей истории (шесть тысяч поколений) человечество пережило три периода, в каждом из которых естественный отбор действовал с неодинаковой силой: период бедствий и катастроф, во время которого люди гибли от холода, голода и насилия, продолжительность жизни была невысокой из-за природных условий, естественный отбор действовал сильно; период болезней (эпидемий), начавшийся несколько тысяч лет назад и до сих пор продолжающий в развивающихся странах, действие естественного отбора сильное; период разрушения организма, при котором человек умирает от старости, действие среды и болезней снижено, так же как и действие естественного отбора снижено. Список литературы
Айала Ф., Кайгер Дж. Современная генетика. Том 3. Москва, Мир, 1988 год Северцов А. С. Основы теории эволюции. Москва, МГУ, 1987 год Программа спецкурсов. Биологический факультет. Спецальность 01.17 – антропология. Москва, МГУ, 2001 год Введение в популяционную и медицинскую генетику. Лекции, А. А. Мовсесян.
В зависимости от количества людей малочисленные популяции делятся на:
Демы. Они включают 1500 - 4000 человек. Они характеризуются:
Низким приростом населения - 20%
Высокой частотой внутригрупповых браков - 80% - 90%
Низким притоком людей из других популяций - 1%-2%.
Изоляты. Они включают до 1500 человек и характеризуются:
Низким приростом населения - 25%
Высокой частотой внутригрупповых браков - более 90%
Низким притоком людей из других популяций - менее 1%.
296. Действие дрейфа генов в популяциях людей и его влияние на генофонды популяций .
Высокая степень репродуктивной изоляции малочисленных человеческих популяций на протяжении многих поколений создавала благоприятные условия для дрейфа генов. Дрейф генов - это изменение частоты аллелей в популяции из-за случайных причин, не обусловленных действием естественного отбора. Значение дрейфа генов: он приводит к изменению частоты аллелей в генофонде популяции. Аллели могут удаляться или закрепляться в генофонде, независимо от того, имеют они адаптивную ценность или нет. Он существенно влияет на генофонд малочисленных популяций. Таким образом, дрейф генов более выражен в малочисленных популяциях.
Естественный отбор против гомозигот в популяциях людей. Пример.
Отбор против гомозигот рассмотрим на примере серповидноклеточной анемии. Есть аллель отвечающий за синтез нормального гемоглобина (HbA), а есть аллель отвечающий за синтез измененного гемоглобина (HbS). Изменённый гемоглобин обладает интересным свойством: в эритроцитах с таким гемоглобином плохо размножается малярийный плазмодий (люди с таким генотипом в 13 раз реже болеют малярией). Таким образом, ребёнок с генотипом HbA/ HbA погибает от малярии, а ребёнок с генотипом HbS/HbS от серповидноклеточной анемии. Если ребенок будет гетерозиготен по этим аллелям (HbA/HbS), то он будет болеть и малярией и серповидноклеточной анемией в лёгкой форме и останется жить. Налицо отбор против гомозигот, он имеет место в районах Африки, Азии, в странах, где распространена малярия.
Естественный отбор против гетерозигот в популяциях людей. Пример.
Отбор против гетерозигот: примером является резус-антиген. Около 80% людей имеют в эритроцитах антиген D. Они резус-положительные Rh+. За синтез антигена отвечает доминантный аллель D. Резус-положительные люди имеют генотип DD или Dd. Резус-отрицательные люди имеют генотип dd. Если резус-отрицательная женщина беременна резус-положительным ребенком, и во время родов эритроциты ребенка попадают в кровь к матери, то у нее в ответ на резус-фактор (антиген) вырабатываются антитела. При второй беременности резус-положительным ребенком, антитела проникают через плаценту в организм ребенка и разрушают его эритроциты. Ребенок может умереть. Т.к. генотип ребенка Dd, отбор направлен против гетерозигот.
Генетический груз в популяциях людей: масштабы, факторы формирования, медицинское значение.
Бремя генетического груза человечества можно оценить, введя понятие летальных эквивалентов. Считают, что число их у отдельных людей колеблется от 3 до 8. Это означает, что общее количество неблагоприятных аллелей, которое есть в генотипе каждого человека, оказывает действие равное действию 3-8 рецессивных аллелей. В результате гомозиготные особи будут умирать до наступления репродуктивного возраста.
Благодаря наличию неблагоприятных аллелей и их сочетаний примерно половина зигот, образующихся в каждом поколении людей, в биологическом плане несостоятельна, т.е. не участвует в передаче генов следующему поколению. Около 15% зачатых организмов гибнет до рождения, 3% - при рождении, 2% - непосредственно после рождения. 3% людей умирает, не достигнув половой зрелости, 20% лиц не вступает в брак и 10% браков бездетны. Благодаря слабому действию естественного отбора в человеческих популяциях и развитию медицины выживает все большее число генетически измененных людей. Они выживают и оставляют такое же генетически измененное потомство. Т.о., величина генетического груза в человеческой популяции постоянно возрастает.
Генетический полиморфизм в популяции людей, его сущность, причины возникновения, масштабы, медицинское значение.
Человек является одним из наиболее полиморфных видов на земле, это наследственное разнообразие проявляется в многообразии фенотипов (цвет кожи, глаз, волос, форма носа и ушной раковины, кожные рисунки пальцев и др.). Этому способствуют мутации и комбинативная изменчивость. Миграции, снижение изоляции, популяционные волны также вносят свой вклад в создание генетического разнообразия популяции людей.
Значение генетического полиморфизма для существования человечества.
Из-за генетического разнообразия заболевания на планете распределены неравномерно.
У разных людей болезни протекают с разной тяжестью.
У людей разная предрасположенность к заболеваниям
Индивидуальные особенности течения патологических процессов.
Различия в реакциях на лечебное воздействие.
Если оценивать генетический полиморфизм в общебиологическом плане, то чем больше генетическое разнообразие людей, тем выше шансы человечества на выживание при контакте с новыми неблагоприятными факторами.
301. Генетический полиморфизм природных популяций, причины создания. Сущность и пример балансированного полиморфизма.
Генетический полиморфизм - это существование в популяции более двух генетически разных форм. Причины полиморфизма: мутации и комбинативная изменчивость. Устанавливается генетический полиморфизм под действием естественного отбора. Балансированный полиморфизм возникает, если естественный отбор больше благоприятствует гетерозиготам, чем гомозиготам. Явление селективного преимущества гетерозигот называют сверхдоминантностью. Пример: в экспериментальной численно равновесной популяции мух, содержащей сначала много мутантов с более темными телами, концентрация последних быстро падала, пока не стабилизировалась на уровне 10 %. Анализ показал, что в созданных условиях гомозиготы по рецессивной мутации и гомозиготы по аллелю дикого типа менее жизнеспособны, чем гетерозиготные мухи.
302. Представить смысловое и математическое выражение закона Харди-Вайнберга, условия его проявления .
■ из поколения в поколение частота генов и генотипов в популяции не изменяется,
если на популяцию не действуют мутации, миграции, естественный отбор, то есть сумма частот генов одного аллеля в данной популяции есть величина постоянная (p+q- 1 или 100%).
■ сумма частот генотипов по одному аллелю в данной популяции есть величина
постоянная, а распределение их соответствует коэффициентам бинома Ньютона второй степени (р + 2pq+ q= 1 или 100%).
Допустим, в популяции присутствуют гены: “А” с частотой “р”, и “а” с частотой “q”.
Тогда: ♂ (p+q)х ♀ (p+q)= (p+q)2=р2 + 2pq+ q2= 1 = 100%
Р – частоту доминантных гомозигот в популяции (АА).
2pq – частоту гетерозигот в популяции (Аа).
q 2 – частоту рецессивных гомозигот в популяции (аа). p+q=1
Условие проявления закона Харди-Вайнберга:
Популяция должна быть достаточно большой.
Должно быть свободное скрещивание особей.
Должна быть равная плодовитость гомозигот и гетерозигот.
Не должны действовать мутации, миграции и естественный отбор.
Популяции, которые отвечают этим условиям, называются идеальными или мен-
делевскими. В природе эти популяции не встречаются.
Что такое дрейф генов. Какое значение имеет это явление.
Дрейф генов - это изменение частоты аллелей в популяции из-за случайных причин, не обусловленных действием естественного отбора. Значение дрейфа генов: он приводит к изменению частоты аллелей в генофонде популяции. Аллели могут удаляться или закрепляться в генофонде, независимо от того, имеют они адаптивную ценность или нет. Он существенно влияет на генофонд малочисленных популяций.
Филогенез систем органов.
Филогенез систем органов.
88. Отбор в пользу и против гетерозигот. Примеры.
Отбор против гетерозигот хорошо виден на примере резус-отрицательных людях. Если резус-фактор матери отрицателен, а отца положителен – происходит эритробластоз плода. Гетерозиготы плохо приспособлены. В популяции происходит гомозиготизация населения. В популяции существует естественный отбор и в пользу гетерозигот. Например, существуют несколько видов гемоглобина. Hb A, Hb S. Они дадут 3 генотипа:
Hb S Hb S – серповидно-клеточная анемия.
В результате точковой мутации гемоглобин теряет растворимость, эритроцит приобретает форму серпа, развивается анемия и весьма вероятен летальный исход (большинство людей не доживают до половой зрелости). Однако в некоторых районах неблагоприятных по малярии было замечено, что гетерозиготы не болеют этим заболеванием и не умирают от нее. В нормальных условиях более приспособлены Hb A Hb A, на равнинах наблюдается гипоксия людей с Hb A Hb S. В неблагоприятных условиях приспособительная способность Hb A Hb A менее 1.
89.Генетический груз и его эволюционное значение
Вывод: беречь окружающую среду.
Генетический груз – часть наследственной изменчивости популяции, определяющая появление менее приспособленных особей, подвергающихся избирательной гибели в результате естественного отбора.
Существует 3 типа генетического груза.
Мутационный.
Сегрегационный.
Субституционный.
Каждый тип генетического груза коррелирует с определенным типом естественного отбора.
Мутационный генетический груз - побочное действие мутационного процесса. Стабилизирующий естественный отбор удаляет вредные мутации из популяции.
Сегрегационный генетический груз – характерен для популяций, использующих преимущество гетерозигот. Удаляются хуже приспособленные гомозиготные особи. Если обе гомозиготы летальны – половина потомков погибает.
Субституционный генетический груз – происходит замена старого аллеля новым. Соответствует движущей форме естественного отбора и переходному полиморфизму.
Впервые генетический груз в популяции человека был определен в 1956г в Северном полушарии и составил 4%. Т.е. 4% детей рождались с наследственной патологией. За последующие годы было введено более миллиона соединений в биосферу (более 6000 ежегодно). Ежедневно – 63000 химических соединений. Растет влияние источников радиоактивного излучения. Структура ДНК нарушается.
Генетическая гибель- гибель организмов, обусловленная естественны отбором, она уменьшает репродуктивный потенциал популяции.
90.Генетический полиморфизм: классификация. Адаптивный потенциал популяции человека
Полиморфизм – существование в единой панмиксной популяции двух и более резко различающихся фенотипов.
Полиморфизм бывает:
Хромосомный;
Переходный;
Сбалансированный.
Генетический полиморфизм наблюдается, когда ген представлен более чем одним аллелем. Пример – системы групп крови.
3 аллеля -А, В, О.
J ª J ª, J ª J ° - А
J ª J в, J в J ° - В
J ª J в - АВ
J ° J ° - О
Генетический полиморфизм широко распространен и лежит в основе наследственной предрасположенности к заболеваниям. Однако болезни наследственных предрасположений проявляются лишь при взаимодействии генов и среды. Условия среды – недостаток или избыток питательных веществ, наличие психогенных факторов, токсических веществ и др.
Генетический полиморфизм создает все условия для протекающей эволюции. При появлении нового фактора в среде популяция способна адаптироваться к новым условиям. Например, устойчивость насекомых к различным видам инсектицидов.
Хромосомный полиморфизм – между особями имеются различия по отдельным хромосомам. Это результат хромосомных аббераций. Есть различия в гетерохроматиновых участках. Если изменения не имеют патологических последствий – хромосомный полиморфизм, характер мутаций – нейтрален.
Переходный полиморфизм – замещение в популяции одного старого аллеля новым, который более полезен в данных условиях. У человека есть ген гаптоглобина - Нр1f, Hp 2fs. Старый аллель - Нр1f, новый - Нр2fs. Нр образует комплекс с гемоглобином и обусловливает слипание эритроцитов в острую фазу заболеваний.
Сбалансированный полиморфизм – возникает, когда ни один из генотипов преимущества не получает, а естественный отбор благоприятствует разнообразию.
Широкий полиморфизм помогает популяции приспосабливаться к условиям среды. У здоровых людей нет противоречия между средой и генотипом, если возникает это противоречие - проявляются болезни наследственного предрасположения.
Есть моногенные и полигенные болезни.
Моногенные болезни наследственного предрасположения – наследственные заболевания, проявляющиеся из-за мутации одного гена или проявляющиеся при действии определенного фактора среды (аутосомно-рецессивные или сцепленные с Х-хромосомой).
Проявляются при воздействии факторов:
Физических;
Химических;
Пищевых;
Загрязнения среды.
Пигментная ксеродерма - веснушчатая кожа особого типа.
Дети не переносят УФ-свет возникают злокачественные опухоли, такие дети умирают от метастаз еще до 15 лет. Не переносят также и гамма-лучей.
Полигенные болезни наследственного происхождения – такие болезни, которые возникают при действии многих факторов (мультифакториальные) и в результате взаимодействия многих генов.
Установить диагноз в таком случае очень сложно, т.к. действует много факторов, и появляется новое качество при взаимодействии факторов.
Генетический полиморфизм человечества: масштабы, факторы формирования. Значение генетического разнообразия в прошлом, настоящем и будущем человечества (медико-биологический и социальный аспекты).
Генетический полиморфизм (наследственное разнообразие) - это сохранение в генофонде популяции различных аллелей одного и того же гена в концентрации, превышающей по наиболее редкой форме 1%. Это разнообразие поддерживается отбором, но создается мутационным процессом. Естественный отбор в этом случае может иметь два механизма: отбор против гомозигот в пользу гетерозигот и отбор против гетерозигот в пользу гомозигот.
В первом случае отбором сохраняются гетерозиготные генотипы популяции и устраняются доминантные и рецессивные гомозиготы. Во втором случае накапливаются в генофонде гомозиготные генотипы и происходит устранение гетерозигот. При действии первого механизма возникает балансированный полиморфизм, при действии второго - адаптационный.
Адаптационный полиморфизм возникает в том случае, когда в различных, но закономерно изменяющихся условиях среды отбор благоприятствует разным генотипам. В человеческих пуляциях это более редкая форма полиморфизма. Наиболее часто проявляется балансированный полиморфизм. Он очень распространен в человеческих популяциях, усиливает гетерозиготизацию, а значит, устойчивость организмов к воздействию факторов среды. Сред-няя степень гетерозиготности в человеческих популяциях составляет 6,7%. Генетическое разнообразие в популяциях человека приводит к фенотипическому разнообразию. Наиболее значительно оно по белковому составу, например по ферментам в генетической системе человека 30% локусов имеют разнообразные гены. У человека имеется около ста полиморфных систем. Значение балансированного полиморфизма заключается в том, что он поддерживает беспредельную генетическую гетерогенность популяции, обеспечивает генетическую индивидуальность каждого человека.
Для медицины изучение балансированного полиморфизма представляет особую важность в связи с тем, что, во-первых, проявляется неравномерность распределения наследственных заболеваний в популяциях; во-вторых, различается степень предрасположенности к болезням; в-третьих, отмечается индивидуальный характер течения болезни и разная ее тяжесть; в-четвертых, имеет место различная ответная реакция на лечебные мероприятия. Отрицательное проявление балансированного полиморфизма проявляется, прежде всего, в наличии генетического груза.
Билет 92.
Макроэволюция. Ее соотношение с микроэволюцией. Формы филогенеза (эволюции групп): филетическая и дивергентная эволюция, конвергентная эволюция и параллелизм. Примеры.
Макроэволюция – это процесс формирования крупных систематических единиц: новых родов, семейств и т.д. Макроэволюция осуществляется на протяжении огромных промежутков времени, и непосредственно изучать ее невозможно. Тем не менее в основе макроэволюции лежат те же движущие силы, что и в основе микроэволюции: наследственная изменчивость, естественный отбор и репродуктивное разобщение.
Понятие макроэволюции. Понятием «макроэволюция» обозначают происхождение надвидовых таксонов (родов, отрядов, классов, типов, отделов). В общем смысле макроэволюцией можно назвать развитие жизни на Земле в целом, включая ее происхождение. Макроэволюционным событием считается также возникновение человека, по многим признакам отличающегося от других биологических видов.
Между микро- и макроэволюцией нельзя провести резкую грань, потому что процесс микроэволюции, первично вызывающий дивергенцию популяций (вплоть до видообразования), продолжается без какого-либо перерыва и на макроэволюци-онном уровне внутри вновь возникших форм.
Отсутствие принципиальных различий в протекании микро- и макроэволюции позволяет рассматривать их как две стороны единого эволюционного процесса и применять для его анализа понятия, разработанные в теории микроэволюции, поскольку макроэволюционные явления (возникновение новых семейств, отрядов и других групп) охватывают десятки миллионов лет и исключают возможность их непосредственного экспериментального исследования.
Среди форм филогенеза выделяют первичные - филетическую эволюцию и дивергенцию, лежащие в основе любых изменений таксонов.
Филетическая эволюция - это изменения, происходящие в одном филогенетическом стволе (без учета всегда возможных дивергентных ответвлений). Без таких изменений не может протекать никакой эволюционный процесс, и поэтому филетическую эволюцию можно считать одной из элементарных форм эволюции. Филетическая эволюция происходит в пределах любой ветви древа жизни: любой вид развивается во времени, и как бы ни были похожи между собой особи вида (разделенные, скажем, несколькими тысячами поколений в неизбежно меняющейся среде), вид в целом должен за это время в чем-то измениться. Это филетическая эволюция на микроэволюционном уровне. Проблемы филетической эволюции на макроэволюционном уровне - изменение во времени близкородственной группы видов.
В «чистом» виде (как эволюция без дивергенции) филетическая эволюция может характеризовать лишь сравнительно короткие периоды эволюционного процесса
Дивергенция - другая первичная форма эволюции таксона. В результате изменения направления отбора в разных условиях происходит дивергенция (расхождение) ветвей древа жизни от единого ствола предков.
Начальные стадии дивергенции можно наблюдать на внутривидовом (микроэволюционном) уровне, на примере возникновения различий по каким-либо признакам в отдельных частях видового населения. Так, дивергенция популяций может приводить к видообразованию
Уже Ч. Дарвин подчеркивал огромную роль дивергенции в процессе развития жизни на Земле. Таков главный путь возникновения органического многообразия и постоянного увеличения «суммы жизни». Механизм дивергентной эволюции основан на действии элементарных эволюционных факторов. В результате изоляции, волн жизни, мутационного процесса и в особенности естественного отбора популяции и группы популяций приобретают и сохраняют в эволюции признаки, все более заметно отличающие их от родительского вида. В какой-то момент эволюции (этот «момент» может длиться много поколений, а для эволюции даже сотни поколений - мгновение) накопившиеся различия окажутся настолько значительными, что приведут к распаду исходного вида на два (и более) новых.
Несмотря на принципиальное сходство процессов дивергенции внутри вида (микроэволюционный уровень) и в группах более крупных, чем вид (макроэволюционный уровень), между ними существует и важное различие, состоящее в том, что на микроэволюционном уровне процесс дивергенции обратим: две разошедшиеся популяции могут легко объединиться путем скрещивания в следующий момент эволюции и существовать вновь как единая популяция. Процессы же дивергенции в макроэволюции необратимы: раз возникший вид не может слиться с прародительским (в ходе филетической эволюции и тот и другой вид неизбежно изменится, и если даже какие-то части этих видов в будущем вступят на путь сетчатой эволюции, или семгенеза, то это не будет возврат к старому.
Дивергенция и филетическая эволюция - основа всех изменений филогенетического древа и первичные формы протекания процесса эволюции любого масштаба в природе.
Самые сложные явления эволюции - это филогенетический параллелизм и филогенетическая конвергенция
Параллелизм - это процесс филетического развития в сходном направлении двух или нескольких генетически близких таксонов. Довольно часто в качестве одной из форм филогенеза называют конвергенцию. Однако конвергентно может возникать только морфофизиологическое сходство по отдельным или нескольким признакам. Образование одного таксона выше уровня вида из двух различных, по-видимому, невозможно.
Очень важно учитывать, что явления направленной эволюции выражаются не только в развитии в одном направлении, но и чаще всего в независимом приобретении организмами ряда общих признаков, отсутствовавших у предков. Если при этом проявляется прямая зависимость специфики приобретаемого признака от функции (например, веретеновидная форма тела у нектонных организмов), то мы говорим о конвергенциях. Если же наряду с функциональными моментами отчетливо проявляется зависимость приобретаемого признака от общих унаследованных особенностей организма, то мы предпочитаем говорить о филогенетических параллелизмах (Татаринов,1983, 1984). Параллелизмы особенно характерны для организмов, связанных относительно тесным родством. Обычно именно этот критерий, измеряемый рангом таксона, кладется в основу различения параллелизмов и конвергенции.
Билет 93.
Макроэволюция. Типы (направления) эволюции групп. Арогенез и ароморфозы. Аллогенез и идиоадаптации. Примеры.
В зависимости от того, изменяется ли уровень организации в эволюционирующих группах, выделяют два основных типа эволюции: аллогенез и арогенез.
При аллогенезе у всех представителей данной группы сохраняются без изменения основные черты строения и функционирования систем органов, благодаря чему уровень организации их остается прежним. Аллогенная эволюция происходит в пределах одной адаптивной зоны - совокупности экологических ниш, различающихся в деталях, но сходных по общему направлению действия основных средовых факторов на организм данного типа. Интенсивное заселение конкретной адаптивной зоны достигается благодаря возникновению у организмов идиоадаптаций - локальных морфофизиологических приспособлений к определенным условиям существования. Пример аллогенеза с приобретением идиоадаптаций к разнообразным условиям обитания в отряде насекомоядных млекопитающих
Арогенез - такое направление эволюции, при котором у некоторых групп внутри более крупного таксона появляются новые морфофизиологические особенности, приводящие к повышению уровня их организации. Эти новые прогрессивные черты организации называют ароморфозами. Ароморфозы позволяют организмам заселять принципиально новые, более сложные адаптивные зоны. Так, арогенез ранних земноводных был обеспечен появлением у них таких основных ароморфозов, как пятипалые конечности наземного типа, легкие и два круга кровообращения с трехкамерным сердцем. Завоевание адаптивной зоны с более сложными для жизни условиями (наземной по сравнению с водной, воздушной по сравнению с наземной) сопровождается активным расселением в ней организмов с появлением у них локальных идиоадаптаций к различным экологическим нишам.
Таким образом, периоды арогенной эволюции группы могут сменяться периодами аллогенеза, когда в результате возникающих идиоадаптаций новая адаптивная зона заселяется и используется наиболее эффективно. Если в ходе филогенеза организмы осваивают более 49
Билет 94.
Макроэволюция. Биологический прогресс и биологический регресс, их основные критерии. Эмпирические правила эволюции групп. Примеры.
Прогресс и его роль в эволюции. На протяжении всей истории живой природы ее развитие осуществляется от более простого к более сложному, от менее совершенного к более совершенному, т.е. эволюция носит прогрессивный характер. Таким образом, общий путь развития живой природы - от простого к сложному, от примитивного к более совершенному. Именно этот путь развития живой природы и обозначают термином «прогресс». Однако всегда закономерно возникает вопрос: почему же в современной фауне и флоре одновременно с высокоорганизованными существуют низкоорганизованные формы? Когда подобная проблема встала перед Ж.Б. Ламарком, он вынужден был прийти к признанию постоянного самозарождения простых организмов из неорганической материи. Ч. Дарвин же считал, что существование высших и низших форм не представляет затруднений для объяснения, так как естественный отбор, или выживание наиболее приспособленных, не предполагает обязательного прогрессивного развития - он только дает преимущество тем изменениям, которые благоприятны для обладающего ими существа в сложных условиях жизни. А если от этого нет никакой пользы, то естественный отбор или не будет вовсе совершенствовать эти формы, или усовершенствует их в очень слабой степени, так что они сохранятся на бесконечные времена на их современной низкой ступени организации.
Процесс эволюции идет непрерывно в направлении максимального приспособления живых организмов к условиям окружающей среды (т. е. происходит возрастание приспособленности потомков по сравнению с предками). Такое возрастание приспособленности организмов к окружающей среде А.Н. Северцов назвал биологическим прогрессом. Постоянное возрастание приспособленности организмов обеспечивает увеличение численности, более широкое распространение данного вида (или группы видов) в пространстве и разделение на подчиненные группы.
Критериями биологического прогресса являются:
увеличение численности особей;
расширение ареала;
прогрессивная дифференциация - увеличение числа систематичес ких групп, составляющих данный таксон.
Эволюционный смысл выделенных критериев заключается в следующем. Возникновение новых приспособлений снижает элиминацию особей, в результате средний уровень численности вида возрастает. Стойкое повышение численности потомков по сравнению с предками приводит к увеличению плотности населения, что, в свою очередь, через обострение внутривидовой конкуренции вызывает расширение ареала; этому же способствует и возрастание приспособленности. Расширение ареала приводит к тому, что вид при расселении сталкивается с новыми факторами среды, к которым необходимо приспосабливаться. Так происходит дифференциация вида, усиливается дивергенция, что ведет к увеличению дочерних таксонов. Таким образом, биологический прогресс - это наиболее общий путь биологической эволюции.
В работах по теории эволюции иногда встречается термин «морфофи-зиологический прогресс». Под морфофизиологическим прогрессом понимают усложнение и совершенствование организации живых организмов.
Регресс и его роль в эволюции. Биологический регресс - явление, противоположное биологическому прогрессу. Он характеризуется снижением численности особей вследствие превышения смертности над рождаемостью, сужением или разрушением целостности ареала, постепенным или быстрым уменьшением видового многообразия группы. Биологический регресс может привести вид к вымиранию. Общая причина биологического регресса - отставание темпов эволюции группы от скорости изменения внешней среды. Эволюционные факторы действуют непрерывно, в результате чего происходит совершенствование приспособлений к изменяющимся условиям среды. Однако когда условия изменяются очень резко (очень часто из-за непродуманной деятельности человека), виды не успевают сформировать соответствующие приспособления. Это приводит к сокращению численности видов, сужению их ареалов, угрозе вымирания. В состоянии биологического регресса находятся многие виды. Среди животных это, например, крупные млекопитающие, такие как уссурийский тигр, гепард, белый медведь, среди растений - гинкговые, представленные в современной флоре одним видом - гинкго двулопастным.
Происхождение и развитие крупных групп организмов (типов, отделов, классов) называется макроэволюцией. Развитие живой природы от более простых форм к более сложным называется прогрессом. Выделяют биологический и морфофизиологический прогресс. Явление, противоположное прогрессу, называется регрессом. Биологический регресс может привести к вымиранию группы в целом или большей части ее видов.
Отбор в пользу гетерозигот, когда обе гомозиготы имеют пониженную по сравнению с гетерозиготой приспособленность, часто называют гетерозисом, или сверхдоминированием. Этот тип отбора существенно отличается от рассмотренного выше направленного отбора: сверхдоминирования приводит к созданию устойчивого полиморфного равновесия.
Рассмотрим изменения генотипических частот за одно поколение отбора при сверхдоминировании. Все рассуждения представлены в таблице 3.
В первых двух строчках приведены приспособленности и частоты генотипов в некотором поколении. Вклад в следующее поколение равен произведению частоты на приспособленность (третья строка). Сумма вкладов всех генотипов называется средней приспособленностью и равна
W = p 2 (l - s) + 2pq + q 2 (l - t).
Последняя строка таблицы соответствует частотам генотипов в следующем поколении. Вычислим изменение частоты аллеля за поколение, опустив часть простых алгебраических выкладок.
Популяция приходит в равновесие, когда частоты аллелей перестают изменяться.
Δq = q − (P aa +
При этом для аллеля А имеем
| s |
| t + s |
В отличие от равновесия в нейтральной популяции (в отсутствии отбора), равновесие при сверхдоминировании является устойчивым. Это означает что при отклонении генных частот от равновесного значения популяция возвращается к тому же равновесному уровню. Напомним, что нейтральная популяция при отклонении от положения равновесия за одно поколение переходит в новое равновесное состояние (с другими частотами генотипов).
Детально изученным примером сверхдоминирования в популяциях человека служит серповидноклеточная анемия - болезнь, широко распространенная в некоторых странах Африки и Азии. Анемия обусловлена аномальным строением гемоглобина (форма S) и развивается у людей гомозиготных по аллелю Hb S . Нормальный гемоглобин вырабатывается в присутствии аллеля Нb A в гомо- или гетерозиготном состоянии. Большинство людей с генотипом Hb A Hb S погибает до достижения половозрелости, так что приспособленность этого генотипа близка к нулю. Несмотря на это, частота аллеля достигает довольно высоких значений в ряде регионов, особенно в районах распространения малярийного плазмодия. Причина этого состоит в том, что гетерозиготы Hb A Hb A более устойчивы к малярии, чем гомозиготы Нb A Нb A , то есть обладают селективным преимуществом по сравнению с обеими гомозиготами, у которых смертность от анемии или от малярии выше, чем у гетерозигот.
А) Отбор против гетерозигот.
Отрицательный отбор в свою очередь может быть направленпротив гетерозигот ипротив гомозигот.
В ряде случаев приспособленность гетерозигот может быть ниже приспособленности гомозигот. Такая картина наблюдается у межвидовых и внутривидовых гибридов, хотя здесь и неприменима однолокусная модель. Низкая приспособленность гетерозигот наблюдается и при наличии транслокаций на одной из хромосом и нормальной парной хромосомы. Такие гетерозиготы по хромосомнымтранслокациям часто образуют несбалансированные гаметы с пониженной жизнеспособностью, что снижает приспособленность гетерозигот по сравнению с гомозиготами. Модель невыгодности гетерозигот может быть полезна для контроля численности популяций вредителей.
Польза гетерозиготности: гетерозиготы по гемофилии, альбинизму, серповидноклеточной анемии и т. д. не страдают данными заболеваниями, вследствие их рецессивности. Особи, несущие полиаллельные гены, дают более жизнеспособное потомство с широкой нормой реакции. Гетерозиготность - это "блокатор" вредных рецессивных аллелей.
Примером отрицательного отбора, направленного против гетерозигот, может служить наследование Rh-фактора.Rh-фактор контролируется тремя доминантными тесно сцепленными генами, поэтому их можно условно принять за один.
При браке мужчины, обладающего резус-положительным фактором, и женщины с резус-отрицательным фактором чаще возможно зачатие "резус-положительного" плода.
Антигены плода в период вынашивания в небольшом количестве способны проникать в кровоток матери через плаценту и вызывать образование антител (особенно если есть патология детского места). При первой беременности (иногда и при второй) концентрация их в крови сравнительно невелика и зародыш развивается, не испытывая вредных влияний этих антител.
Б) Отбор против гомозигот.
Во многих случаях происходит лишь частичный отбор против гомозигот. Поэтому относительная приспособленность гомозигот по сравнению с другими генотипами уменьшается лишь отчасти. При многих генетических болезнях человека, например при альбинизме или серповидноклеточной анемии , гомозиготы по рецессивному аллелю могут выжить и дать потомство, хотя и с меньшей вероятностью, чем здоровые индивиды. У дрозофилы, мыши, кукурузы и других генетически исследованных организмов существует множество рецессивных мутаций, которые снижают приспособленность, но не приводят к смертельному исходу.
В) отбор и контротбор.
Контротбор-положительный отбор, направленный против отрицательного отбора (поддержание гомозигот и гетеорозигот)
Генофонд популяций человека является результатом наложения многочисленных и разнонаправленных векторов отбора, обеспечивающего сохранение в каждом поколении сравнительно приспособленных к данным условиям генотипов. При этом с течением времени влияние отбора на генетическую структуру популяций людей снижается в основном благодаря успехам лечебной и профилактической медицины, а также социально-экономическим преобразованиям цивилизации.
Вопрос№ 20 учение о микро-и макроэволюции.
А) Микроэволюция.
Микроэволюция - это распространение в популяциималых изменений в частотахаллелейна протяжении несколькихпоколений; эволюционные изменения на внутривидовом уровне. Такие изменения происходят из-за следующих процессов:мутации,естественный отбор,искусственный отбор,перенос геновидрейф генов. Эти изменения приводят кдивергенциипопуляций внутривида, и, в конечном итоге, квидообразованию.
Под действием элементарных факторов на генофонд популяции происходит изменение частот отдельных генов. Это приводит к элементарному эволюционному явлению - изменению генотипического и фенотипического состава популяции. При длительном однонаправленном воздействии естественного отбора наблюдается дифференциация популяций.
Б) биологическая сущность макроэволюции.
Сущность макроэволюции. Этим понятием обозначают происхождение надвидовых таксонов (родов, отрядов, классов, типов, отделов). В общем смысле макроэволюцией можно назвать развитие жизни на Земле в целом, включая и ее происхождение. Макроэволюционным событием считается также возникновение человека, по многим признакам отличающегося от других биологических видов. Между микро- и макроэволюцией нельзя провести резкую грань, потому что процесс микроэво-
люции, первично вызывающий дивергенцию популяций (вплоть до видообразования), продолжается без какого-либо перерыва и на макроэволюционном уровне внутри вновь возникших форм.
Отсутствие принципиальных различий в протекании микро- и макроэволюционно! о процессов позволяет рассматривать их как две стороны единого эволюционного процесса и применять для анализа всего процесса понятия, разработанные в теории микроэволюции, поскольку макроэволюционные явления охватывают десятки миллионов лет и исключают возможность их непосредственного экспериментального исследования.
Способы осуществления макроэволюции. Макроэволюция может осуществляться несколькими способами. Основной способ -дивергенция - представляет собой независимое образование различных признаков у родственных организмов. В основе дивергенции лежит экологическая дифференциация вида (или группы видов) на самостоятельные ветви. Различия между видами одной группы в процессе эволюции, в силу изменения направления отбора, все более и более углубляются. Но вместе с тем сохраняется и определенная общность признаков морфофизиологической организации. Это свидетельствует о происхождении данной группы от общего родоначального предка. При дивергенции сходство между организмами объясняется общностью их происхождения, аразли-чия - приспособлением к разным условиям среды.
Примером дивергенции форм является возникновение разнообразных по морфофизиологическим особенностям вьюрков от одного или немногих предковых видов на Галапагосских островах. Расхождение внутривидовых форм и видов по разным местообитаниям определяется конкуренцией в борьбе за одинаковые условия, выход из которых и заключается в расселении по разным экологическим нишам.
Еще один способ осуществления макроэволюции -параллелизм (параллельное развитие). Это процесс эволюционного развития в сходном направлении двух или нескольких первоначально дивергировавших групп. Например, палеонтологи очень часто обнаруживают асинхронный параллелизм, т. е. независимое приобретение сходных черт родственными, но живущими в разное время организмами. Примером может служить развитие саблезубости у представителей разных подсемейств кошачьих. С генетической точки зрения параллельная эволюция объясняется общностью генной структуры родственных групп и сходной ее изменчивостью.
В эволюции может наблюдаться также конвергенция (конвергентное развитие) - процесс эволюционного развития двух или более неродственных групп в сходном направлении. Конвергенция обусловлена одинаковой средой обитания, в которую попадают неродственные организмы. Классическим примером конвергентного развития считается возникновение сходных форм тела у акуловых (первичноводные формы), ихтиозавров и китообразных (вторичноводные формы). При конвергентном развитии сходство между неродственными организмами бывает всегда только внешним (эволюционным изменениям в одном направлении подвергаются внешние признаки как результат приспособления к одинаковым условиям среды). По форме тела ихтиозавр похож на акулу и дельфина, но по таким существенным чертам, как строение кожных покровов, черепа, мускулатуры, кровеносной системы, дыхательной и других систем, эти группы позвоночных различны. При конвергентном способе эволюции возникают аналогичные органы.
В) Мегаэволюция. Мегаэвол ю ция , совокупность процессов эволюции живых форм, определяющая формирование крупных таксонов - систематических категорий выше отряда (у животных) и порядка (у растений
Г) Понятие об элементарной эволюц. структуре и материале.